
中国西南地区3种蝇子草属植物的核型报道
The Karyotype Study of Three Species of Silene (Caryophyllaceae) in Southwest China
-
Keywords:
- Silene /
- karyotype /
- polyploidy
-
细胞分类学(Cytotaxonomy)是20世纪初在细胞形态学的基础上发展起来的一门新兴学科,其实质是将染色体的数目、形态结构、行为、DNA含量以及核苷酸重复序列等细胞学资料运用在分类学上,进而探讨和研究类群之间的亲缘关系、分类地位、物种形成机制和进化关系等问题[1-5]。在细胞分类学的基础上,通过与其他学科的交叉融合,还发展出了物种生物学(Biosystematics)、细胞地理学(Cytogeography)等衍生学科[3, 6]。中国西南地区植物类群丰富,且成分复杂,仅云南就具有几乎所有的分布型和亚型,特有现象明显[7];细胞分类学可以解决常规形态分类难以解决的问题,是现代综合分类的重要组成部分[8],对于该地区的植物资源利用及系统分类学具有重要意义。
蝇子草属(Silene L.)为石竹科(Caryophyllaceae)一年或多年生草本植物,全球约400种。中国有112种2亚种17变种,广布长江流域和北部各省区,以西北和西南地区较多[9]。横断山地区共有49种18个种下单位[10]。在国外,蝇子草属植物的染色体数目有较多研究,且大部分种已开展细胞学研究[11-12]。但是在西南横断山地区,截至目前有染色体报道的蝇子草属植物只有7种,分别是细蝇子草(S. gracilicaulis,2n=2x=24=22m+2sm)、垫状蝇子草(S. davidii,2n=4x=48=48m)、尼泊尔蝇子草(S. nepalensis,2n=2x=24=16m+8sm)、变黑蝇子草(S. nigrescens,2n=4x=48=40m+8sm)、内蒙古女娄菜(S. orientalimongolica,2n=2x=24=24m)、糙叶蝇子草(S. trachyphylla,2n=2x=24=8m+6sm+10st)和腺毛蝇子草(S. yetii,2n=6x=72=64m+8sm)[13]。对于该地区染色体的报道十分有限,需要更多的数据补充。
本研究对3种蝇子草属植物进行核型的首次报道,分别为(1)喜马拉雅蝇子草(S. himalayensis),分布于中国河北、湖北、陕西、四川、云南和西藏等省区,以及印度锡金地区,生于海拔2 000~5 000 m的灌丛间或高山草甸;(2)喇嘛蝇子草(S. lamarum)为多年生草本,分布于四川(盐源、西昌、雷波、木里)和云南(德钦、中甸),生于海拔3 400~4 000 m的高山草地或灌丛中;(3)墨脱蝇子草(S. namlaensis),为多年生草本,高30~40 cm,叶片狭椭圆形或披针形,花单生或2~3朵,微俯垂,后期直立蒴果卵形;花期8—9月。墨脱蝇子草分布于中国西藏(林芝、波密、墨脱、加查、措那、聂木拉),生长在海拔3 600~4 500 m的高山草地[9]。
1. 材料与方法
1.1 材料
本试验供试材料的采集具体地点见表1。凭证标本存放在中国科学院昆明植物研究所标本馆(KUN)及西南林业大学林学院标本室,永久装片保存于云南师范大学生命科学学院植物细胞学实验室。
表 1 蝇子草属3种植物材料的来源信息Table 1. The original information of three species of Silene种名species 采集地location 海拔/m altitude 经纬度position 标本号sample No. 喜马拉雅蝇子草 S. himalayensis 四川稻城Daocheng, Sichuan 4 420 E100°16′42.25′′, N28°52′54.96′′ MS17-689 喇嘛蝇子草 S. lamarum 云南昆明Kunming, Yunnan 4 101 E102°55′40.09′′, N26°09′21.48′′ MSC-058 墨脱蝇子草 S. namlaensis 西藏林芝Linzhi, Tibet 4 578 E94°38′50.47′′, N29°36′59.08′′ Tsui-645 1.2 方法
把野外收集的种子放置于4 ℃冰箱,然后将种子置于固体培养基,24 ℃恒温培养箱萌发,待其根尖长到1~2 cm时取下,用0.003 mol/L的8-羟基喹啉常温下避光处理3 h,之后用蒸馏水清洗2~3次,用卡诺氏固定液(无水乙醇∶冰乙酸= 3∶1)在4 ℃冰箱固定24 h,用蒸馏水清洗之后,加入1 mol/L的HCl,在60 ℃ 水浴锅中解离10~15 min,随后清洗,用改良苯酚品红染液染色24 h,使用常规压片法镜检观察,挑选处于分裂中期的分散良好的细胞拍照并制作永久装片。
染色体核型分析标准按照李懋学等[14]的标准,按照ARANO[15]的方法计算核型不对称系数:As.K =(染色体组长臂总和/染色体组总长度和)× 100%;参考PASZKO[16]、PERUZZI等[17]和ASTUTI等[18]对核型参数使用的建议,得出相关核型参数及核型公式;最后取5~6个染色体分散良好的细胞照片进行数据分析。
2. 结果与分析
2.1 喜马拉雅蝇子草[S. himalayensis (Rohrb.) Majumdar]
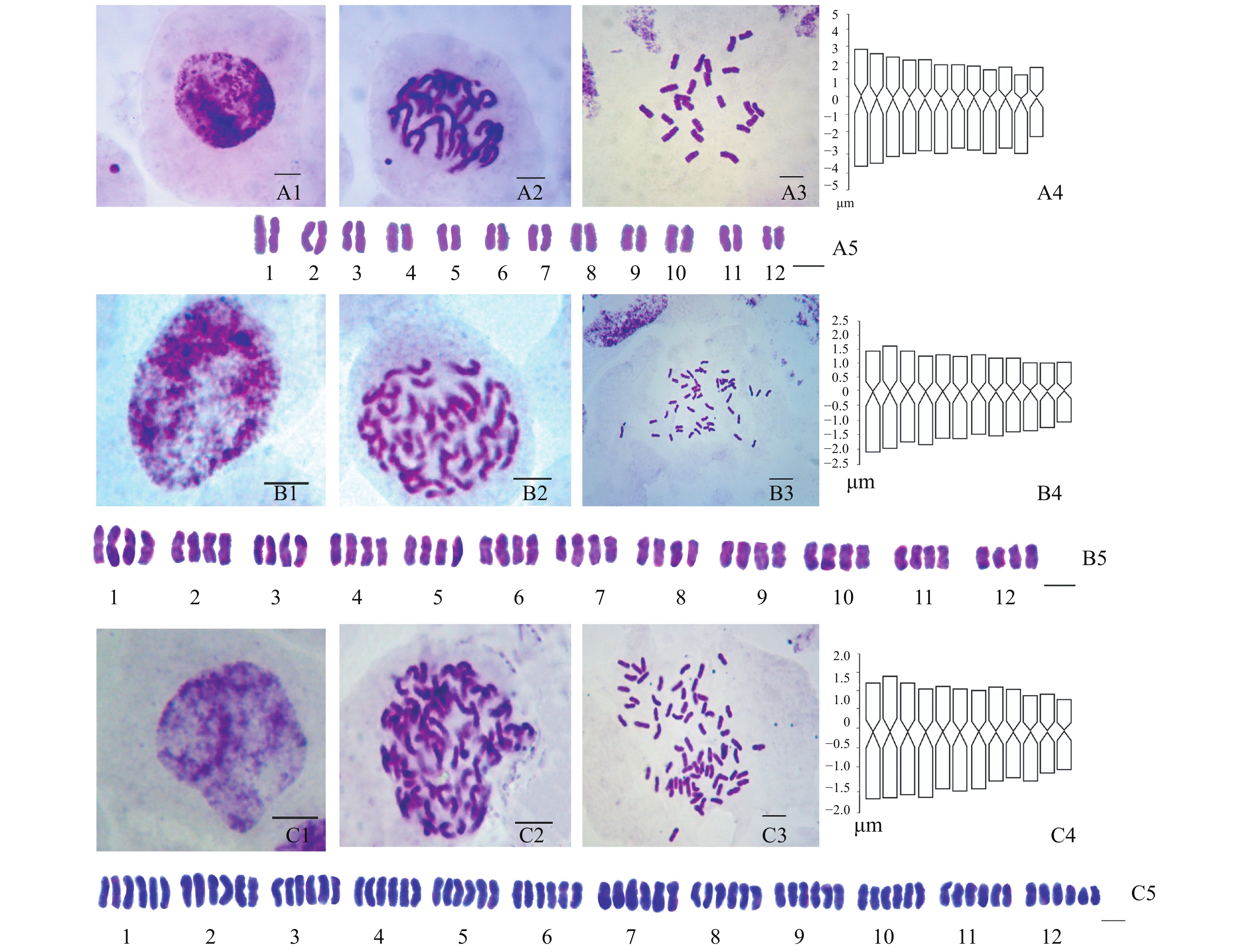
喜马拉雅蝇子草染色体数目为2n=48,核型公式为2n=4x=48m,为四倍体,核型类型为1A型;染色体长度范围为3.78~2.24 μm,臂比范围为1.51~1.26 μm,核型不对称指数(As.K)为56.85%,臂比>2∶1的为0,染色体内不对称程度(MCA)为13.68。染色体参数如表2、3所示,染色体核型模式图如图1所示。
表 2 蝇子草属3个种的染色体参数Table 2. Chromosome parameters of three species of Silene种名 species 染色体序号 chromosome No. 长臂 L 短臂 S 总长 L+S 臂比 L/S 染色体类型 chromosome type 喜马拉雅蝇子草
S. himalayensis1 2.15 1.41 3.55 1.54 m 2 1.96 1.59 3.55 1.25 m 3 1.75 1.41 3.16 1.24 m 4 1.86 1.25 3.11 1.50 m 5 1.66 1.29 2.94 1.29 m 6 1.67 1.26 2.93 1.33 m 7 1.48 1.27 2.75 1.17 m 8 1.56 1.19 2.74 1.30 m 9 1.44 1.19 2.62 1.21 m 10 1.46 1.03 2.49 1.41 m 11 1.40 1.02 2.41 1.38 m 12 1.30 1.03 2.33 1.27 m 喇嘛蝇子草
S. lamarum1 3.40 2.91 6.31 1.17 m 2 3.41 2.45 5.86 1.39 m 3 3.03 2.32 5.35 1.31 m 4 2.95 2.23 5.18 1.32 m 5 2.71 2.20 4.91 1.23 m 6 3.01 1.84 4.84 1.64 m 7 2.45 2.15 4.61 1.14 m 8 2.82 1.84 4.66 1.53 m 9 2.97 1.40 4.37 2.12 m 10 2.53 1.80 4.33 1.41 m 11 2.96 1.11 4.07 2.67 m 12 2.33 1.75 4.08 1.33 sm 墨脱蝇子草
S. namlaensis1 1.67 1.24 2.90 1.34 m 2 1.63 1.36 3.00 1.20 m 3 1.56 1.26 2.81 1.24 m 4 1.62 1.03 2.66 1.58 m 5 1.43 1.17 2.59 1.22 m 6 1.50 1.07 2.57 1.40 m 7 1.43 1.00 2.43 1.42 m 8 1.29 1.11 2.40 1.16 m 9 1.24 1.02 2.27 1.22 m 10 1.30 0.9 2.19 1.44 m 11 1.14 0.97 1.12 1.17 m 12 1.1 0.81 1.91 1.36 m 注:L. 长臂;S. 短臂;L+S. 染色体长度;L/S. 臂比;m. 中部着丝粒区;sm.近中部着丝粒区。
Note: L. long arm; S. short arm; L+S. total length; L/S. arms ratio; m. median centeomere chromosome; sm. submedian centromere chromosome.表 3 蝇子草属3个种的染色体核型参数Table 3. Karyotype and chromosomal data of three species of Silene种名species 染色体数目/基数/倍性
chromosome No./basic No./polyploidy核型karyotype CVCL CVCI AI THL MCA As.K/% 核型公式
karyotype formula喜马拉雅蝇子草
S. himalayensis48/12/4x 1A 14.16 5.06 0.72 34.58 13.68 56.94 2n=4x=48m 喇嘛蝇子草
S. lamarum24/12/2x 2A 14.21 14.18 2.01 58.57 18.85 59.02 2n=2x=19m+5sm 墨脱蝇子草
S. namlaensis72/12/6x 1A 13.39 5.79 0.76 29.85 13.23 56.65 2n=6x=72m 注:CVCL. 染色体长度变异系数; CVCI. 着丝粒指数变异系数; AI. 染色体不对称指数; THL. 核型单倍体染色体总长; MCA. 染色体内不对称程度; As.K. 核型不对称指数。
Note:CVCL. coefficient of variation of chromosome length; CVCI. coefficient of variation of centromere index; AI. chromosome asymmetry index; THL. total chromosome length of karyotype haploid; MCA. degree of intrachromosomal asymmetry; As.K. karyotype asymmetry index.![]() 图 1 3种蝇子草的核型细胞及模式图注:A1~A5. 喇嘛蝇子草;B1~B5. 喜马拉雅蝇子草;C1~C5. 墨脱蝇子草;1~5分别代表间期核、前期核、中期核、核型模式图和核型图。Figure 1. The metaphase chromosomes and idiograms of SileneNote: A1-A5. S. lamarum; B1-B5. S. himalayensis; C1-C5. S. namlaensis; 1-5 interphase nucleus, prophase nucleus, metaphase nucleus, idiogram and karyogram.
图 1 3种蝇子草的核型细胞及模式图注:A1~A5. 喇嘛蝇子草;B1~B5. 喜马拉雅蝇子草;C1~C5. 墨脱蝇子草;1~5分别代表间期核、前期核、中期核、核型模式图和核型图。Figure 1. The metaphase chromosomes and idiograms of SileneNote: A1-A5. S. lamarum; B1-B5. S. himalayensis; C1-C5. S. namlaensis; 1-5 interphase nucleus, prophase nucleus, metaphase nucleus, idiogram and karyogram.2.2 喇嘛蝇子草(S. lamarum C. Y. Wu)
喇嘛蝇子草染色体数目为2n=24,为二倍体,核型公式为2n=2x=19 m+5 sm,核型类型为2A型;染色体长度范围为3.4~2.33 μm,臂比范围为1.14~2.67 μm,核型不对称指数(As.K)为59.02%,臂比>2∶1的占16.7%,染色体内不对称程度(MCA)为18.85。染色体参数如表2、3所示,染色体核型模式图如图1所示。
2.3 墨脱蝇子草[S. namlaensis (Marquand) Bocquet]
墨脱蝇子草染色体数目为2n=72,核型公式为2n=6x=72,核型类型为1A型,染色体长度范围为2.9~1.91 μm,臂比范围为1.58~1.16 μm, 核型不对称指数(As.K)为56.65%,染色体内不对称程度(MCA)为13.23。染色体参数如表2、3所示,染色体核型模式图如图1所示。
3. 讨论
本研究首次报道了蝇子草属3个种的核型及染色体数目,3个种的间期核形态都相似,染色质比较均匀地分布在细胞核内,周围有染色较浅的染色质丝和纤维状结构,根据TANAKA[19]的间期核划分标准为扩散型(diffuse type);有丝分裂前期核染色体上着色较深且比较均匀,隶属于中间型前期核(interstitial type)。有丝分裂中期的染色体数目为:喇嘛蝇子草(S. lamarum)为24条,是二倍体;喜马拉雅蝇子草(S. himalayensis)为48条,是四倍体;墨脱蝇子草(S. namlaensis)为72条,是六倍体。3个种的染色体基数都是12,但是在倍性上存在不同,分别是二倍、四倍和六倍,这可能是由于它们生长的地理位置导致的倍性变化,物种为了适应恶劣的环境,改变自身的倍性,产生多倍化现象,这也是物种分化变异的主要来源之一。在本研究中,墨脱蝇子草生长的海拔总体较高,其次是喜马拉雅蝇子草、喇嘛蝇子草,从这个角度来看,随着海拔的升高,倍性也在增加,但是由于试验数据有限,更普遍的结论需要更进一步的试验验证。
从核型类型来看,喜马拉雅蝇子草和墨脱蝇子草核型都属于1A型,喇嘛蝇子草则为2A型,根据STEBBINS[20]的理论,在生物的进化过程中,染色体核型是由对称性向非对称性演化的。理论认为:核型对称程度越高的生物,其染色体变异越小,进化程度也越低;而非对称程度越高的生物,其染色体变异越大,进化程度越高。按照这一理论及本试验结果可推测:在系统演化上,喜马拉雅蝇子草和墨脱蝇子草属于进化程度相对较低的种类,而喇嘛蝇子草的染色体则属于进化程度比较高的类型。本研究结果可为蝇子草属染色体提供更多的数据补充,并为蝇子草属的系统分类学及探讨多倍化现象、多倍化物种形成等研究提供有价值的参考。
致谢:感谢西南林业大学林学院徐波、中国科学院昆明植物研究所马祥光和孙文光提供试验种子以及云南师范大学生命科学学院苏富明、饶培瑜和胡晶晶在试验上的帮助。
-
![]()
图 1 3种蝇子草的核型细胞及模式图
注:A1~A5. 喇嘛蝇子草;B1~B5. 喜马拉雅蝇子草;C1~C5. 墨脱蝇子草;1~5分别代表间期核、前期核、中期核、核型模式图和核型图。
Figure 1. The metaphase chromosomes and idiograms of Silene
Note: A1-A5. S. lamarum; B1-B5. S. himalayensis; C1-C5. S. namlaensis; 1-5 interphase nucleus, prophase nucleus, metaphase nucleus, idiogram and karyogram.
表 1 蝇子草属3种植物材料的来源信息
Table 1 The original information of three species of Silene
种名species 采集地location 海拔/m altitude 经纬度position 标本号sample No. 喜马拉雅蝇子草 S. himalayensis 四川稻城Daocheng, Sichuan 4 420 E100°16′42.25′′, N28°52′54.96′′ MS17-689 喇嘛蝇子草 S. lamarum 云南昆明Kunming, Yunnan 4 101 E102°55′40.09′′, N26°09′21.48′′ MSC-058 墨脱蝇子草 S. namlaensis 西藏林芝Linzhi, Tibet 4 578 E94°38′50.47′′, N29°36′59.08′′ Tsui-645  下载: 导出CSV
下载: 导出CSV
表 2 蝇子草属3个种的染色体参数
Table 2 Chromosome parameters of three species of Silene
种名 species 染色体序号 chromosome No. 长臂 L 短臂 S 总长 L+S 臂比 L/S 染色体类型 chromosome type 喜马拉雅蝇子草
S. himalayensis1 2.15 1.41 3.55 1.54 m 2 1.96 1.59 3.55 1.25 m 3 1.75 1.41 3.16 1.24 m 4 1.86 1.25 3.11 1.50 m 5 1.66 1.29 2.94 1.29 m 6 1.67 1.26 2.93 1.33 m 7 1.48 1.27 2.75 1.17 m 8 1.56 1.19 2.74 1.30 m 9 1.44 1.19 2.62 1.21 m 10 1.46 1.03 2.49 1.41 m 11 1.40 1.02 2.41 1.38 m 12 1.30 1.03 2.33 1.27 m 喇嘛蝇子草
S. lamarum1 3.40 2.91 6.31 1.17 m 2 3.41 2.45 5.86 1.39 m 3 3.03 2.32 5.35 1.31 m 4 2.95 2.23 5.18 1.32 m 5 2.71 2.20 4.91 1.23 m 6 3.01 1.84 4.84 1.64 m 7 2.45 2.15 4.61 1.14 m 8 2.82 1.84 4.66 1.53 m 9 2.97 1.40 4.37 2.12 m 10 2.53 1.80 4.33 1.41 m 11 2.96 1.11 4.07 2.67 m 12 2.33 1.75 4.08 1.33 sm 墨脱蝇子草
S. namlaensis1 1.67 1.24 2.90 1.34 m 2 1.63 1.36 3.00 1.20 m 3 1.56 1.26 2.81 1.24 m 4 1.62 1.03 2.66 1.58 m 5 1.43 1.17 2.59 1.22 m 6 1.50 1.07 2.57 1.40 m 7 1.43 1.00 2.43 1.42 m 8 1.29 1.11 2.40 1.16 m 9 1.24 1.02 2.27 1.22 m 10 1.30 0.9 2.19 1.44 m 11 1.14 0.97 1.12 1.17 m 12 1.1 0.81 1.91 1.36 m 注:L. 长臂;S. 短臂;L+S. 染色体长度;L/S. 臂比;m. 中部着丝粒区;sm.近中部着丝粒区。
Note: L. long arm; S. short arm; L+S. total length; L/S. arms ratio; m. median centeomere chromosome; sm. submedian centromere chromosome.
下载: 导出CSV
表 3 蝇子草属3个种的染色体核型参数
Table 3 Karyotype and chromosomal data of three species of Silene
种名species 染色体数目/基数/倍性
chromosome No./basic No./polyploidy核型karyotype CVCL CVCI AI THL MCA As.K/% 核型公式
karyotype formula喜马拉雅蝇子草
S. himalayensis48/12/4x 1A 14.16 5.06 0.72 34.58 13.68 56.94 2n=4x=48m 喇嘛蝇子草
S. lamarum24/12/2x 2A 14.21 14.18 2.01 58.57 18.85 59.02 2n=2x=19m+5sm 墨脱蝇子草
S. namlaensis72/12/6x 1A 13.39 5.79 0.76 29.85 13.23 56.65 2n=6x=72m 注:CVCL. 染色体长度变异系数; CVCI. 着丝粒指数变异系数; AI. 染色体不对称指数; THL. 核型单倍体染色体总长; MCA. 染色体内不对称程度; As.K. 核型不对称指数。
Note:CVCL. coefficient of variation of chromosome length; CVCI. coefficient of variation of centromere index; AI. chromosome asymmetry index; THL. total chromosome length of karyotype haploid; MCA. degree of intrachromosomal asymmetry; As.K. karyotype asymmetry index.
下载: 导出CSV
-
[1] 杨纯瑜, 王徽勤. 中国植物细胞分类学的回顾与展望[J]. 植物学通报, 1985, 20(6): 1. [2] 洪德元. 植物细胞分类学[M]. 北京: 科学出版社, 1990. [3] DAVIS P, HEYWOOD V. Principles of angiosperms taxonomy[J]. Oliver & Boyd, Edinburgh and London, 1965, 53(2): 212A.
[4] 吴甘霖. 核型分析在细胞分类学中的应用[J]. 生物学杂志, 2006, 23(1): 39. DOI: 10.3969/j.issn.2095-1736.2006.01.013. [5] 孙文光. 豆科山羊豆族鱼鳔槐属的分子系统学研究[D]. 昆明: 云南师范大学, 2016. [6] SWANSON C P. Chromosome Botany. C.D. Darlington[J]. Quarterly Review of Biology, 1956, 31(4): 300. DOI: 10.1086/401514.
[7] 彭华. 中国西南地区植物资源与农业生物多样性[J]. 云南植物研究, 2001, 23(S1): 29. [8] 巩红冬. 青藏高原东缘二十五种风毛菊属植物的核型研究及分类学意义[D]. 兰州: 西北师范大学, 2007. [9] 王文采. 横断山区维管植物(下册) [M]. 北京: 科学出版社, 1994. [10] BARI E A. Cytological studies in the genus Silene L[J]. New Phytologist, 1973, 72(4): 833. DOI: 10.1111/j.1469-8137.1973.tb02059.x.
[11] SHEIDAI M, EFTEKHARIAN R, GHOLIPOOR A, et al. Population diversity and polyploidy incidence in 3 silene species: a cytologcal approach[J]. Cytologia, 2011, 76(4): 395. DOI: 10.1508/cytologia.76.395.
[12] 李志敏, 何文洁. 横断山区石竹科细蝇子草的染色体数目及核型报道[J]. 云南植物研究, 2007, 29(4): 439. DOI: 10.3969/j.issn.2095-0845.2007.04.011. [13] LUO D, LIU D, XU B, et al. A karyological study of six species of Silene L. (Caryophyllaceae) from the Hengduan Mountains, SW China[J]. Caryologia, 2011, 64(1): 10. DOI: 10.1080/00087114.2011.10589759.
[14] 李懋学, 陈瑞阳. 关于植物核型分析的标准化问题[J]. 武汉植物学研究, 1985, 3(4): 297. [15] ARANO H. Cytological studies in subfamily Carduoideae (Compositae) of Japan XIV[J]. Shokubutsugaku Zosshi, 1963, 76(906): 459. DOI: 10.15281/jplantres1887.76.459.
[16] PASZKO B. A critical review and a new proposal of karyotype asymmetry indices[J]. Plant Systematics and Evolution, 2006, 258(1/2): 39. DOI: 10.1007/s00606-005-0389-2.
[17] PERUZZI L, ALTINORDU F. A proposal for a multivariate quantitative approach to infer karyological relationships among taxa[J]. Comparative Cytogenetics, 2014, 8(4): 337. DOI: 10.3897/compcytogen.v8i4.8564.
[18] ASTUTI G, ROMA-MARZIO F, PERUZZI L. Traditional karyomorphological studies: can they still provide a solid basis in plant systematics?[J]. Flora Mediterranea, 2017, 27: 91.
[19] TANAKA R. Types of resting nuclei in Orchidaceae[J]. Shokubutsugaku Zosshi, 1971, 84: 118. DOI: 10.15281/jplantres1887.84.118.
[20] STEBBINS G L. Chromosomal evolution in higher plants[M]. New York: Addison Wesley, 1971.
计量
- 文章访问数: 2440
- PDF下载量: 14